A group of gas vesicles. Aerotopes appear as brownish, refractive bodies within cells. There may be numerous aerotopes distributed all throughout the cell contents, or there may be only one or a few, localized to a specific area within the cell. When aerotopes are distributed throughout the cell contents, the cell may look dark brown and grainy/mottled (for example, Microcystis). These structures influence the buoyancy of cyanobacteria cells, and are found mostly in planktic species; many of the genera very common in surface blooms (Aphanizomenon, Dolichospermum, Microcystis, Woronichinia) have aerotopes distributed throughout their cells.
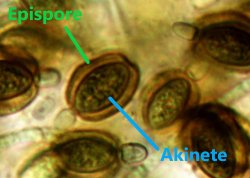
Specialized cell type. Dormant cells, occurring in some filamentous taxa (for example, Anabaena), which allow cyanobacteria to survive harsh environmental conditions and to begin growing when conditions improve; they also aid in dispersal. Akinetes arise from one or more vegetative cells. They have thickened cell walls and contain reserves of the materials needed for sustained survival and eventual germination into new vegetative cells. They are typically larger than vegetative cells.
When used generally (as “terminal cell” or “end cell”), “apical cell” refers to the cell at the very end (either end) of a trichome. When describing heteropolar trichomes, the term “apical” is more specific, because there is also a “basal” end; if the trichome is attached to the substrate, the apical end is the free (unattached) end.
Apical end of a trichome with especially elongated and narrowed (only rarely not narrowed) cells. Almost exclusively a feature of heteropolar types, but may develop in some isopolar types as well.
Apoheterocytic
Describes a certain type/pattern of akinete development: akinetes start developing roughly halfway between two distant heterocytes (“distant” meaning not directly adjacent to the akinete). They develop successively in rows, proceeding in both directions towards the heterocytes. All vegetative cells may may change into akinetes (only in some taxa).
In heteropolar trichomes, the basal end or base is the end by which the trichome is attached to the substrate. However, such trichomes are not always attached to the substrate. For instance: in younger, more spherical colonies of Rivularia, the basal ends of trichomes are oriented towards the center of the colony. A basal heterocyte–meaning a terminal heterocyte at the basal end–is often associated with heteropolarity. The cells and sheath at the base of a trichome may be morphologically differentiated.
A morphological feature of some filamentous cyanobacteria; a thickened, cap-like structure formed on the outside wall of the terminal cell. The calyptra is formed by remnants of either a sheath or a necridic cell.
Capitate
(Describing a terminal cell) Cap-like or knob-like, abruptly swollen/widened.
The outer layer of an akinete cell wall. May be widened, textured, and/or yellowish to orange-brownish in color.
False branching
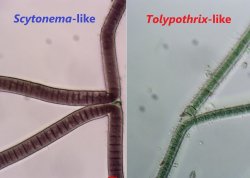
Also referred to as “pseudobranching.” False branches are the result of trichome breakage within a sheath, followed by the continued growth of one or both ends of the trichome. Unlike in true branching, false branches are not formed due to changes in the plane of cell division; the plane of cell division remains perpendicular to the long axis of the trichome. Trichome breakage may occur at heterocytes, or may occur between vegetative cells (facilitated by necridic cell formation). There are two types of false branching: Scytonema-like false branching (also referred to as double or geminate false branching) and Tolypothrix-like false branching (may also be referred to as single false branches). Scytonema-like false branching entails the formation of two morphologically identical paired branches, both diverging away from the original axis of the trichome. Tolypothrix-like false branching entails the formation of a single branch that diverges away from the main trichome axis.
May refer to: a single trichome with its surrounding sheath (filament=trichome+sheath); multiple trichomes, arranged in parallel, with their common surrounding sheath; a single trichome, if that trichome does not have a sheath.
Hollow cylindrical compartments, with walls composed of protein. An aerotope is a group of gas vesicles clustered together, arranged parallel to one another.
An inclusion within a cell, visible with a light microscope. Types of granules include carotenoids (pigment type), and storage (storing chemical reserves) granules. Granules may be distributed throughout the cell, or concentrated in a specific region of the cell.
Specialized cell type. Heterocytes contain an enzyme called nitrogenase, enabling fixation of atmospheric molecular nitrogen. Heterocyte production tends to decrease as the amount of biologically available nitrogen in the environment increases. Though the name for this cell type was once “heterocyst,” the proper name (revised in order to reflect their actual function) is indeed “heterocyte.”
Describes a cell, trichome, filament, or thallus, in which one end is morphologically different from the other. The basal end is often attached to a substrate, while the apical end is free (unattached).
Singular: hormogonium. Short segments of trichomes which separate from the (original) trichome, for the purposes of trichome reproduction and dispersal. Hormogonia separate from the ends of trichomes and the ends of branches. They may also separate from a position between vegetative cells in the trichome (rather than at the end of the trichome or at a branch tip) following the disintegration of the trichome by the formation of heterocytes or necridic cells. The cells within a hormogonium may look different from the original trichome, and may contain aerotopes and/or storage granules. Hormogonia are commonly motile, sometimes intensely so.
(Of a cell or structure; within this site, commonly used to describe heterocytes) Between cells within a trichome, as opposed to at one end of a trichome.
Used here in the context of heterocyte position. Apical at the end of a short branch (true branching); or in a multiseriate trichome, forming after lengthwise division of a marginal cell.
(Of a trichome) Heterocytes and/or akinetes develop at roughly regular distances along the length of the trichome.
Moniliform
(Of a trichome) Composed of roughly spherical to oval cells, giving the trichome a bead-like appearance.
Multiseriate
(Of a trichome) With cells capable of dividing in a direction or directions other than just perpendicular to the trichome’s long axis, resulting in more than one row of cells (roughly parallel to one another).
Describes a certain pattern of akinete development: akinetes develop right next to, or slightly distant from, heterocytes, and then arise successively, proceeding away from the heterocyte. May develop in rows of up to 8.
Mucilaginous structure surrounding cells or trichomes. Morphology varies: some sheaths are firm and distinct, while others are fine and diffluent; they may have clearly visible layers (lamellate), and they may be pigmented (most commonly with a yellowish-brown color).
Terminal cell
The cell at the very end (not specific; refers to either end) of a trichome. Also referred to as “end cell.”
In the context of cyanobacteria, this term refers to the whole “body” of the cyanobacterial growth; commonly used to describe macroscopic colonies. For instance, if a cyanobacterium grows as a mat-like layer of many filaments covering the substrate to which they are attached, one might say that “the thallus is in the form of a mat.”
Trichome
A uniseriate or multiseriate row of connected cells; these cells form one physiological unit, in the sense that there is physiological transport between the cells within a trichome. Typically, the term “trichome” refers just to the row(s) of cells, and does not include any enveloping sheath that may be present.
True branches are branches that remain physiologically connected with the main trichome; they are not the result of trichome breakage within the trichome’s sheath. These branches are formed through the following process: a cell within a trichome divides in a plane that is not perpendicular to the trichome’s long axis, then one or both of the resulting cells change their direction of division, and subsequent divisions result in growth that is perpendicular or oblique to the original trichome. True branching only occurs in heterocytous cyanobacteria, though many heterocytous types do not display true branching.
(Of a trichome) If obligately uniseriate: with cells capable of dividing only in the plane that perpendicular to the trichome’s long axis, resulting in only one row of cells. Some multiseriate forms may have portions of the trichome or branches that have just one row of cells, and some true-branching types may be uniseriate (though cell division in other planes occurs, hence the true branching).
Johansen, J. R., & Komárek, J. (2015). Coccoid Cyanobacteria. In J. D. Wehr, R. G. Sheath, & J. P. Kociolek (Eds.), Freshwater Algae of North America: Ecology and Classification (2nd ed., pp. 80-81). Waltham, MA: Elsevier.
Komárek, J. (2013). Cyanoprokaryota-3. Teil/Part 3: Heterocytous Genera (J. R. Johansen, Ed.). In B. Büdel, G. Gärtner, L. Krienitz, & M. Schagerl (Eds.), Süßwasserflora von Mitteleuropa (Vol. 19/3, pp. 7-65). Heidelberg, Germany: Springer Spektrum.
Komárek, J. & Anagnostidis, K. (2008). Cyanoprokaryota-1. Teil/Part 1: Chroococcales. In H. Ettl, G. Gärtner, H. Heynig, & D. Mollenhauer (Eds.), Süßwasserflora von Mitteleuropa (Vol. 19/1, p. 30-32). Heidelberg, Germany: Spektrum.
Wehr, J. D., Sheath, R. G., & Kociolek, J. P. (2015). Freshwater algae of North America: ecology and classification (2nd ed.). London: Elsevier Academic Press.